Studies on Salt Tolerance of Transgenic Sweetpotato Which Harbors Two Genes Expressing CuZn Superoxide Dismutase and Ascorbate Peroxidase with the Stress-inducible SWPA2 Promoter 
2. Environmental Biotechnology Research Center, Korea Research Institute of Bioscience and Biotechnology, Daejeon, 305-806, Korea
 Author
Author  Correspondence author
Correspondence author
Plant Gene and Trait, 2012, Vol. 3, No. 2 doi: 10.5376/pgt.2012.03.0002
Received: 19 Sep., 2011 Accepted: 28 Nov., 2011 Published: 07 Dec., 2011
Wang et al., 2011, Studies on Salt Tolerance of Transgenic Sweetpotato which Harbors Two Genes Expressing CuZn Superoxide Dismutase and Ascorbate Peroxidase with the Stress-inducible SWPA2 Promoter, Plant Gene and Trait, Vol.3, No.2 6-12 (doi: 10.5376/pgt.2012.03.0002)
In this study, some physiological indexes of leaves in transgenic sweetpotato (Ipomoea batatas L. cv. Yulmi), which harbors two genes CuZn superoxide dismutase (CuZnSOD) and ascorbate peroxidase (APX) genes, with the stress-inducible SWPA2 promoter were evaluated under different concentrations of NaCl treatment. The results showed that physiological indexes were no remarkable differences without NaCl stress between the transgenic sweetpotato (TS) and the non-transformed sweet potato (NS). The activity of superoxide dismutase (SOD), ascorbate peroxidease (APX), peroxidase (POD) and catalase (CAT) in leaves of TS was always higher than NS under the same NaCl stress, respectively, and when the concentration NaCl with 100 mmol/L, the enzyme activities was the most significant difference especially. On the other hand, the root length of TS was longer than that of NS. And the decline range of chlorophyll and malonaldehyde (MDA) content in leaves of TS was lower than that of NS. All these results indicated that transgenic sweetpotato had the resistance to salt tolerance. Therefore, there would be a great significance in efficiently utilizing saline land and alleviating the energy crisis by developing and planting transgenic sweetpotato plants with salt tolerance.
There are a lot of wasteland in the coastal areas, the north and northwest of China because of the soil salinization. Sweetpotato (Ipomoea batatas (L.) Lam.) as an important food, feed, industrial raw material and new energy source crops, is a strong applicability crop. Therefore, there would be a great significance in efficiently utilizing saline land and alleviating the energy crisis by developing and planting transgenic sweetpotato plants with salt tolerance.
Under environment stresses, the antioxidant defense system of plants will play a coordinating role to effectively scavenge the superoxide radicals, H2O2, singlet oxygen and hydroxyl free radicals, aviod lipid peroxidation, and prevent the cell membrane from harm. SOD, CAT, POD and APX play an important role in removing excess reactive oxygen species (ROS) of cellular, among them, two key ROS detoxification enzymes in the chloroplast are SOD and APX. SOD is the first substance in common defense response against abiotic stresses and catalyzes the dismutation of two molecules of the superoxide anion radical into oxygen and hydrogen peroxide, and APX reduces hydrogen peroxide to water by utilizing ascorbate as an electron donor (Mittler, 2002). Recently, SOD genes have been overexpressed in plants to improve their tolerance to environmental stresses (Yu et al., 1999; Kwon et al., 2002). Liu et al (2003) used transgenic potato plants as the materials to study the changes of active oxygen metabolism and protective enzyme at different NaCl concentrations stress, the results showed that the transgenic potato with CuZn-SOD gene had a strong antioxidant capacity and a potential ability in salt tolerance. The previous work demonstrated that transgenic plant expressing the genes of APX improved its tolerance to oxidative stress (Kornyeyev et al., 2001). Transgenic tobacco plants expressing both CuZnSOD and APX provide stronger protection to methyl viologen (MV)-induced oxidative stress than transgenic tobacco plants expressing CuZnSOD, MnSOD or APX, which was relative with SOD and APX scavenging ROS at the same time in different path (Kwon et al., 2002). There were many reports indicated that transgenic sweetpotato plants expressing the genes of CuZnSOD and APX in chloroplasts improved its tolerance to drought, chilling, high temperature and sulfur dioxide stress responses (Lim et al., 2007; Li and Deng, 2007).
The foreign gene cannot express without high-effective expression promoter. The constitutive promoter CaMV was used in the most transgenic researches; however, this kind of constitutive promoter drives the gene to express regardless of time and space. For example, Kasuga et al (1999) reported that with the help of CaMV 35S promoter, it can drive the expression of the DREB1A gene related to drought resistance resulted in severe growth retardation under normal growing conditions. The stress-inducible promoter has the transcriptional activity because of the signal stimulation, comparing to tissue and organ-specific promoter, it can drive the target gene up-regulation expression only after the plants was stimulation by signal, so that it not only does not result in waste of resources in plants, but also improve resistance of plant. Kim et al (2003) reported that the isolation of a strong oxidative stress-inducible promoter of POD (SWPA2) from sweetpotato and then characterized that it was more effective to drive resistant gene expression in transgenic tobacco plants. The application of SWPA2 enhanced tolerance to environmental stresses of transgenic potato and sweetpotato plants (Tang et al., 2006; Lim et al., 2007; Ahmad et al., 2008). Therefore, the stress-inducible promoter might be useful for the development of stress-tolerant transgenic plants.
In this Study, some physiological indexes of leaves in transgenic sweetpotato (Ipomoea batatas L. cv. Yulmi), which harbors two genes CuZn superoxide dismutase (CuZnSOD) and ascorbate peroxidase (APX) genes with the stress-inducible SWPA2 promoter were evaluated under different concentrations of NaCl treatment, which would provide a basis for breeding stress-tolerant sweetpotato plants.
1 Results and Analysis
1.1 PCR analysis of transgenic plants
PCR analyses were performed to determine integration of SOD, APX, and neomycin phosphotransferase â…¡ (NPTâ…¡) genes in the genomic DNA of transgenic plants. The result showed that the predicted 458 bp, 752 bp and 700 bp internal fragments for the SOD, APX, and NPTâ…¡, respectively, were amplified in the transgenic plant, indicating that those genes were integrated into the genome (Figure 1).
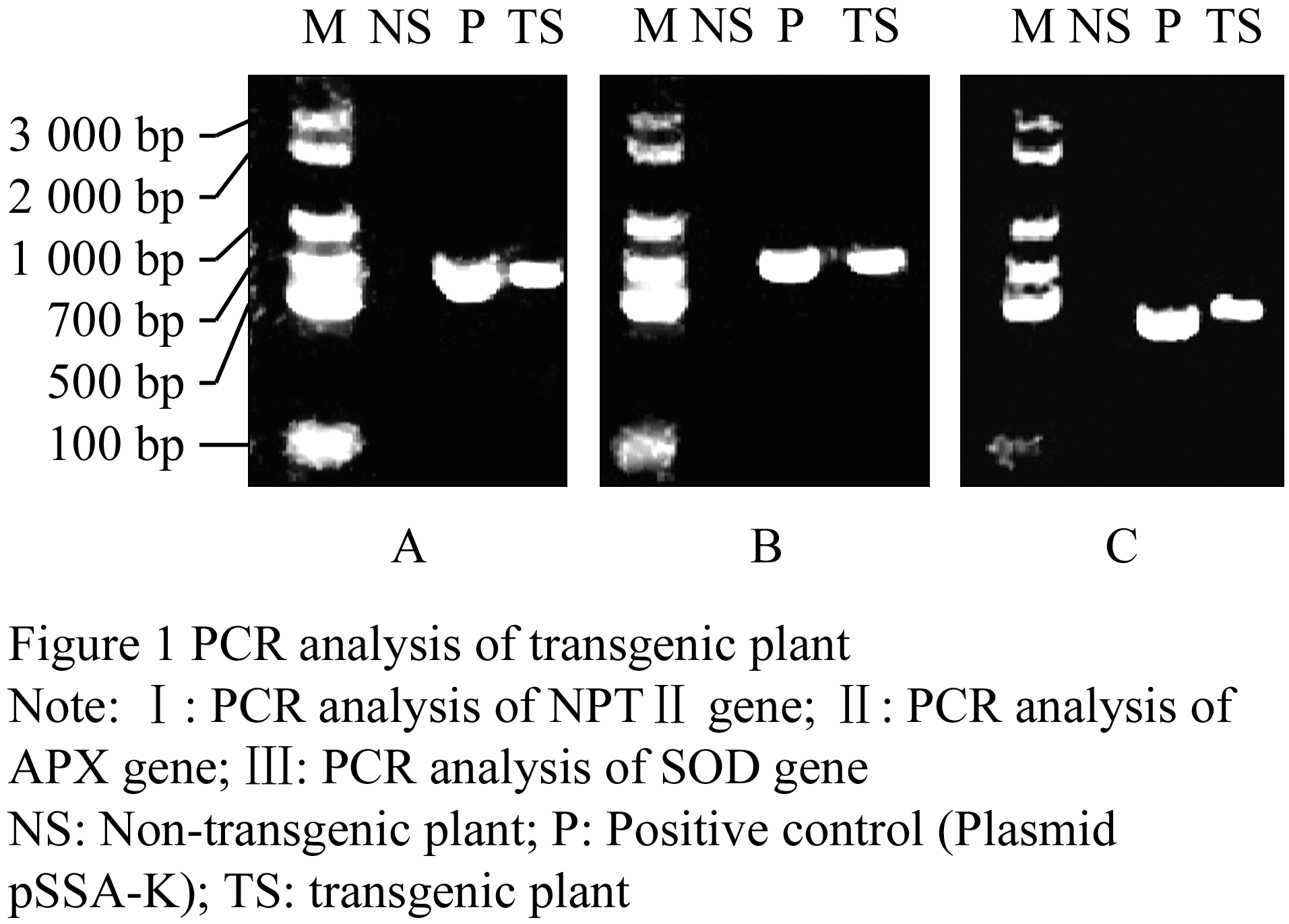
Figure 1 PCR analysis of transgenic plant
1.2 Anti-oxidation enzyme assays
The enzyme activities of SOD, APX, CAT and POD showed approximately the same trend in transgenic sweetpotato under NaCl stress (Figure 2). The levels of SOD, APX, CAT and POD activities in leaves of TS were always higher than those in NS under the same NaCl stress, which indicated that excessive expression of SOD-APX gene were also raised POD and CAT activities of the antioxidant enzyme system, thus strengthen the ability to withstand attack of superoxide radical, etc. Physiological indexes were no remarkable differences without NaCl stress between TS and NS, which was concerned with the stress-inducible promoter that did not start the expression of foreign genes. With the increase of the concentration of the Nacl, the enzyme activity in TS improved quickly, and reached the peak value in 100 mmol/L NaCl concentration. We observed remarkable differences under 100 mmol/L NaCl concentration between TS and NS (P<0.01). In the case of the APX activity, this exhibited a significant difference (P<1%) from NS at 100 mmol/L NaCl, approximately 4-fold higher than that without NaCl treatment and 7-fold higher than the NS at the 100 mmol/L NaCl concentration (Figure 2). When salt stress was in 150 mmol/L NaCl concentration levels, the enzyme activity decreased in TS and NS, because cell plasmid membrane was destructed seriously and antioxidant enzymes were inactivates. By contrast to TS, the enzyme activities of NS decrease sharply, even increased few or decreased below the 100 mmol/L NaCl concentration level, SOD activity decreased under the 50 mmol/L NaCl concentration level. These showed that transgenic sweetpotato might efficiently tolerate the salt stress, because excessive expression of SOD-APX gene with the stress-inducible promoter.
.png) Figure 2 SOD, APX, CAT and POD activities in sweet potato leaves under NaCl stress |
1.3 MDA (Malondialdehyde) content
The MDA is one of the final products of the cell membrane damage because of free radicals attack. The MDA content positively correlated with NaCl concentration. There was no remarkable difference without NaCl stress between TS and NS, however, there were remarkable differences under NaCl stress between TS and NS (P<0.01 and P<0.01; Figure 3). The MDA content of TS was lowest under the 100 mmol/L NaCl concentration level, resulting from high enzyme activities which cleaned extra MDA, thereby reduced the damage of the cell membrane. When salt stress was in 150 mmol/L NaCl concentration levels, the MDA content increased in TS and NS, because cell plasmid membrane was destructed and the enzymes activities declined.
.png) Figure 3 MDA content in sweet potato leaves under NaCl stress |
1.4 Chlorophyll content
The chlorophyll content of the leaf reduced due to NaCl stress (Rao and Rao, 1981). There was no remarkable difference without NaCl stress between TS and NS. The decline range of the chlorophyll in the leaves of TS was smaller than in the NS with NaCl stress. There were remarkable differences under the 100 mmol/L and 150 mmol/L NaCl stress between TS and NS (P <0.01) (Table 1).
Table 1 The chlorophyll content of leaf under the NaCl stress |
Ke and Pan (1999) reported salt stress could lead to disintegration of chloroplast fine structure and make pigment protein complex instability, so the chlorophyll was destroyed, and inevitably reduced the absorption of light energy. The Table 1 showed that the chloroplasts of NS were destroyed seriously, which of TS were also a certain degree of damage under 150 mmol/L NaCl concentration level.
1.5 The root length
Under normal conditions, the root of the TS and NS could grow very well. With the increase of the NaCl concentration, the root growth was restrained, but the root length of TS was longer than the NS. There were remarkable differences under the 100 mmol/L and 150 mmol/L NaCl stress between TS and NS (P<0.01; Figure 4). In addition, the author also found the more fibrous root of the TS grew than the NS in 50 mmol/L NaCl and 100 mmol/L NaCl concentration level. This meant that the transgenic plants could be better to resist salt harm.
.png) Figure 4 The root length under NaCl stress under NaCl stress |
2 Discussion
In the long course of evolution, the plant formed the perfect and complex protection system including enzymes and non-enzymatic antioxidant systems to cope with oxidative stress (Du et al., 2001). The SOD and APX are two key enzymes in the ROS scavenging system (Perl et al., 1993; Badawi et al., 2004), which could clear out active oxygen on different path (Asada, 1999). Transgenic tobacco plants expressing both CuZnSOD and APX provide stronger protection to methyl viologen (MV)-induced oxidative stress than transgenic tobacco plants expressing CuZnSOD or APX (Kwon, 2002). In this study, we also confirmed that transgenic sweetpotato plants expressing both CuZnSOD and APX improved the salt resistance.
In view of the foregoing results, with NaCl concentration increasing, the activity of SOD, APX, CAT and POD in sweetpotato leaves was increased firstly and then decreased, while in general condition, the activity of these four enzymes in transgenic sweetpotato was higher than no-transgenic sweetpotato. When antioxidant enzyme activities increased, H2O2 decreased in cell, which increased the stability of membranes and CO2 fixation because the enzymes of the Calvin cycle within chloroplasts are extremely sensitive to H2O2. When the H2O2 level rose to a certain degree to trigger foreign gene over-expression, consequently higher antioxidant enzyme activities of TS occurred at 100 mmol/L NaCl, especially APX activity had a substantial increase at 100 mmol/L NaCl. The different increasing value of APX activity and other antioxidant enzyme activities suggest that APX might be responsible for the fine modulation of ROIs signaling during stress, and that of CAT might be responsible for removal of excess ROIs (Mittler, 2002). Further intensification of the NaCl stress caused decreases of antioxidant enzyme activities in both sweetpotato plants, because cytoplasmic membrane was destructed seriously and antioxidant enzymes were inactivated. But the degrees of decline were moderate in transgenic sweetpotato and sharp in no-transgenic sweetpotato.
The MDA content and the decline range of the chlorophyll of TS were lower than that of in the NS under NaCl stress. These results were agreed to the results of the root length. These correlated with improvement of the enzymes activities. The MDA content and the decline range of the chlorophyll of TS increased a little with increasing a little of the enzyme activities under the 50 mmol/L NaCl concentration level. This might due to the response time to NaCl stress signal. With the enzyme activities improved quickly under the 100 mmol/L NaCl concentration level, The MDA content of TS decreased a little and the decline range of the chlorophyll of TS did not increased so much, consequently here it became evident that ROS cleaning system is a complex balance process and the high enzyme activities could not cleaned out the whole of the MDA.
All of the above results indicate that the tolerant improvement of transgenic sweetpotato to salt stress is accompanied with the enhancement of antioxidative capacity. With the development of research, transgenic plants with salt tolerance would be applied in the saline land and improve the ecological environment.
3 Materials and Methods
3.1 Materials
Transgenic sweetpotato plants with CuZnSOD and APX genes and sweetpotato plants cv. Yulmi as control (Lim et al., 2007) grew under normal growth conditions until 6~7 leaf stage. And then, their stems with 5 leaves were treated with solutions containing 0 (as control), 50 mmol/L, 100 mmol/L and 150 mmol/L NaCl and maintained for 5 days with 3 replicates, each replicate consisting of 5 plants.
3.2 Analysis of PCR
The genomic DNA of the TS and NS leaves was extracted according to modified CTAB method (Li et al., 2007). The specific primers for the PCR analysis were designed according to the sequences of NPTⅡ, SOD and APX. A 750 bp product approximately was amplified with the NPTⅡ primer (5'-GAGGCTATTCGGCTATGACTG-3', 5'-ATCGGGAGCGGCGATACCGTA-3'); a 752 bp product was amplified with the APX primer (5'-ATGGGAAAATCTTACCCAACTGTTA-3', 5'-TTAGGCTTCAGCAAATCCAAGCTC-3'), as well as the SOD primer (5'-ATGGTGAAGGCTGAAGCTGTTCTT-3', 5'-CTATCCTCGCAAACCAATACCG-3') with a 458 bp product. PCR amplification reactions were initially incubated at 94℃ for 5 min, followed by 30 cycles at 94℃ for 1 min, 62℃ (NPTⅡ, SOD) or 60℃ (APX) for 1 min, and 72℃ for 1 min, at last 72℃ for 5 min. The reaction products were analyzed by 1.2% agarose gel electrophoresis.
3.3 Physiological indexes assays
The third fully expanded leaf from the top of the NS and TS plants was collected to assay for SOD, APX, CAT, POD activities and MDA content, respectively. Leaf samples (1 g) were homogenized in ice cold 0.1 M phosphate buffer (pH=7.8 for SOD and CAT extraction, pH=7.0 for APX, POD and MDA extraction) containing 1% PVP (polyvinyl pyrrolidone) with pre-chilled pestle and mortar. Each homogenate was transferred to centrifuge tubes and was centrifuged at 4℃ for 15 min at 12 000 ×g. The supernatant was used for enzyme activity assay.
SOD activity was measured by recording the decrease in absorbance of superoxidenitro blue tetrazolium complex by the enzyme (Beauchamp and Fridovich, 1971). Absorbance was recorded at 560 nm and one unit of SOD activity was defined as that which inhibited 50% of the reaction rate under these conditions. APX activity was measured according to Shen et al (1996) by monitoring the rate of ascorbate oxidation at 290 nm. 1 unit enzyme activity was computed by enzyme amount decreasing 0.1 in absorbance per minter. CAT activity was measured according to Zeng et al (1991). 1 unit enzyme activity was computed by enzyme amount decreasing 0.1 in absorbance per minute. POD activity was measured using methyl catechol. Absorbance was recorded at 470 nm for 0 min, 1 min, 2 min and 3 min. 1 unit enzyme activity was computed by enzyme amount increasing 0.1 in absorbance per minute. MDA content was measured using thiobarbituric acid reagent according to Zhao et al (1994).
3.4 The chlorophyll content
The chlorophyll content was measured by Chlorophyll Content Meter (USA, Opti-Sciences, CM-200). The same location of the third fully expanded leaf from the top of the NS and TS plants was collected to assay with 3 replicates, each replicate consisting of 3 leaves.
3.5 The root length
The root length was measured with 3 replicates, each replicate consisting of 20 roots, after growing in different NaCl concentration levels for 5 days.
3.6 Data analysis
DPS software was used for analysis the variance and significant differences in tests.
Authors' contributions
WX conceived the overall study, performed the experiment designs and drafted the manuscript. GXM and TZH took part in the experiment; LQ performed part of the data analysis. KSS and MDF performed the experiment designs. MDF read the manuscript and revised it. All authors had read and consent the final text.
Acknowledgements
This study was supported by Chinese National Programs for High Technology Research and Development (“863”program, Code# 2009AA10Z102) and Chinese National Science and Technology Support Program (2009BADA7B03).
References
Ahmad R., Kim M.D., Back K.H., Kim H.S., Lee H.S., Kwon S.Y., Murata N., Chung W.I., and Kwak S.S., 2008, Stress-induced expression of choline oxidase in potato plant chloroplasts confers enhanced tolerance to oxidative, salt, and drought stresses, Plant Cell Reports, 27(4): 687-698
http://dx.doi.org/10.1007/s00299-007-0479-4
Asada K., 1999, The water-water cycle in chloroplasts: scavenging of active oxygens and dissipation of excess photons, Annual Review of Plant Physiology and Plant Molecular Biology, 50: 601-639
http://dx.doi.org/10.1146/annurev.arplant.50.1.601
Badawi G.H., Kawano N., Yamauchi Y., Shimada E., Sasaki R., Kubo A., and Tanaka K., 2004, Over-expression of ascorbate peroxidase in tobacco chloroplasts enhances the tolerance to salt stress and water deficit, Physiologia Plantarum, 121(2): 231-238
http://dx.doi.org/10.1111/j.0031-9317.2004.00308.x
Beauchamp C., and Fridovich I., 1971, Superoxide dismutase: improved assays and an assay applicable to acrylamide gels, Analytical Biochemistry, 44(1): 276-287
http://dx.doi.org/10.1016/0003-2697(71)90370-8
Du X.M., Yin W.X., Zhao Y.X., and Zhang H., 2001, The production and scavenging of reactive oxygen species in plants, Shengwu Gongcheng Xuebao (Chinese Journal of Biotechnology), 17(2): 121-125
Kasuga M., Liu Q., Miura S., Yamaguchi-Shinozaki K., and Shinozaki K., 1999, Improving plant drought, salt, and freezing tolerance by gene transfer of a single stress-inducible transcription factor, Nature Biotechnology, 17: 287-291
http://dx.doi.org/10.1038/7036
Ke Y.Q., and Pan T.G., 1999, Effects of salt stress on the ultrastructure of chloroplast and the activities of some protective enzymes in leaves of sweet potato, Zhiwu Shengli Xuebao (Acta hytophysiologica Sinica), 25(3): 229-233 (in Chinese)
Kim K.Y., Kwon S.Y., Lee H.S., Hur Y., Bang J.W., and Kwak S.S., 2003, A novel oxidative stress-inducible peroxidase promoter from sweetpotato: molecular cloning and characterization in transgenic tobacco plants and cultured cells, Plant Molecular Biology, 51(6): 831-838
http://dx.doi.org/10.1023/A:1023045218815
Kornyeyev D., Logan B.A., Payton P., Allen R.D., and Holaday A.S., 2001, Enhanced photochemical light utilization and decreased chilling-induced photoinhibition of photosystem â…¡ in cotton over-expressing genes encoding chloroplast-targeted antioxidant enzymes, Physiologia Plantarum, 113(3): 323-331
http://dx.doi.org/10.1034/j.1399-3054.2001.1130304.x
Kwon S.Y., Lee H.S., Kim J.S., Cho K.Y., Allen R.D., and Kwak S.S., 2002, Enhanced tolerance of transgenic tobacco plants expressing both superoxide dismutase and ascorbate peroxidase in chloroplasts against methyl viologen-mediated oxidative stress, Plant Cell and Environment, 25: 873-882
http://dx.doi.org/10.1046/j.1365-3040.2002.00870.x
Li Q., Qie Q., Liu Q.C., Wang X., Ma D.F., Zhai H., and Wang Y.P., 2007, An efficient and rapid method for sweetpotato genomic DNA extraction, Fenzi Zhiwu Yuzhong (Molecular Plant Breeding), 5(5): 743-746 (in Chinese)
Li Y.M., and Deng X.P., 2007, Antioxidative system characteristics of sweet potato transferred both Cu/Zn superoxide dismutase and ascorbate peroxidase gene under water stress and rewatering, Nongye Gongcheng Kexue (Chinese Agricultural Science Bulletin), 23(6): 616-621 (in Chinese)
Lim S., Kim Y.H., Kim S.H., Kwon S.k., Lee H.S., Kim J.S., Cho K.Y., Paek K.Y., and Kwak S.S., 2007, Enhanced tolerance of transgenic sweetpotato plants that express both CuZnSOD and APX in chloroplasts to methyl viologen-mediated oxidative stress and chilling, Molecular Breeding, 19(3): 227-239
http://dx.doi.org/10.1007/s11032-006-9051-0
Liu X.P., Zhao Y.C., Huang D.E., Li W.D., and Yuan Q.S., 2003, Metabolism of active oxygen and change of cell defense enzyme in potato transfered with CuZn-SOD gene under NaCl stress, Zhiwu Baohu (Plant Protection), 29(3): 21-24 (in Chinese)
Mittler R., 2002, Oxidative stress, antioxidants and stress tolerance, Trends Plant Science, 7(9): 405-410
http://dx.doi.org/10.1016/S1360-1385(02)02312-9
Perl A., Perl-Treves R., Galili S., Aviv D., Shalgi E., Malkin S., and Galun E., 1993, Enhanced oxidative stress defence in transgenic potato expressing Cu,Zn superoxide dismutase, Theoretical and Applied Genetics, 85(5): 568-576
http://dx.doi.org/10.1007/BF00220915
Rao G.G., and Rao G.R., 1981, Pigment composition & chlorophyllase activity in pigeon pea (Cajanus indicus Spreng) & Gingelley (Sesamum indicum L.) under NaCl salinity, Indian Journal of Experimental Biology, 19: 768-770
Shen W.B., Xu L.L., Ye M.B., and Zhang R.X., 1996, Study on determination of ASP activity, Zhiwu Shengli Tongxun (Plant Physiology Communications), 32(3): 203-205 (in Chinese)
Tang L., Kwon S.Y., Kim S.H., Kim J.S., Choi J.S., Cho K.Y., Sung C.K., Kwak S.S., and Lee H.S., 2006, Enhanced tolerance of transgenic potato plants expressing both superoxide dismutase and ascorbate peroxidase in chloroplasts against oxidative stress and high temperature, Plant Cell Reports, 25(12): 1380-1386
http://dx.doi.org/10.1007/s00299-006-0199-1
Yu Q., Osborne L.D., and Rengel Z., 1999, Increased tolerance to Mn deficiency in transgenic tobacco overproducing superoxide dismutase, Annals of Botany, 84(4): 543-547
http://dx.doi.org/10.1006/anbo.1999.0951
Zeng S.X., Wang Y.R., and Liu H.X., 1991, Some enzymatic reactions related to chlorophyll degradation in cucumber cotyledons under chilling in the light, Zhiwu Shengli Xuebao (Acta Phytophysiologica Sinica), 17(2): 177-182 (in Chinese)
Zhao S.J., Xu C.C., Zou Q., and Meng Q.W., 1994, Improvements of method for measurement of malondialdehyde in plant tissues, Zhiwu Shengli Tongxun (Plant Physiology Communications), 30(3): 207-210 (in Chinese)



. PDF(695KB)
. FPDF(win)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Xin Wang
. Xiaoming Guo
. Qiang Li
. Zhonghou Tang
. Sangsoo Kwak
. Daifu Ma
Related articles
. Transgenic sweetpotato
. CuZnSOD
. APX
. Salt tolerance
Tools
. Email to a friend
. Post a comment